Divulgação científica na área de Fisiologia do Treinamento e Nutrição Esportiva
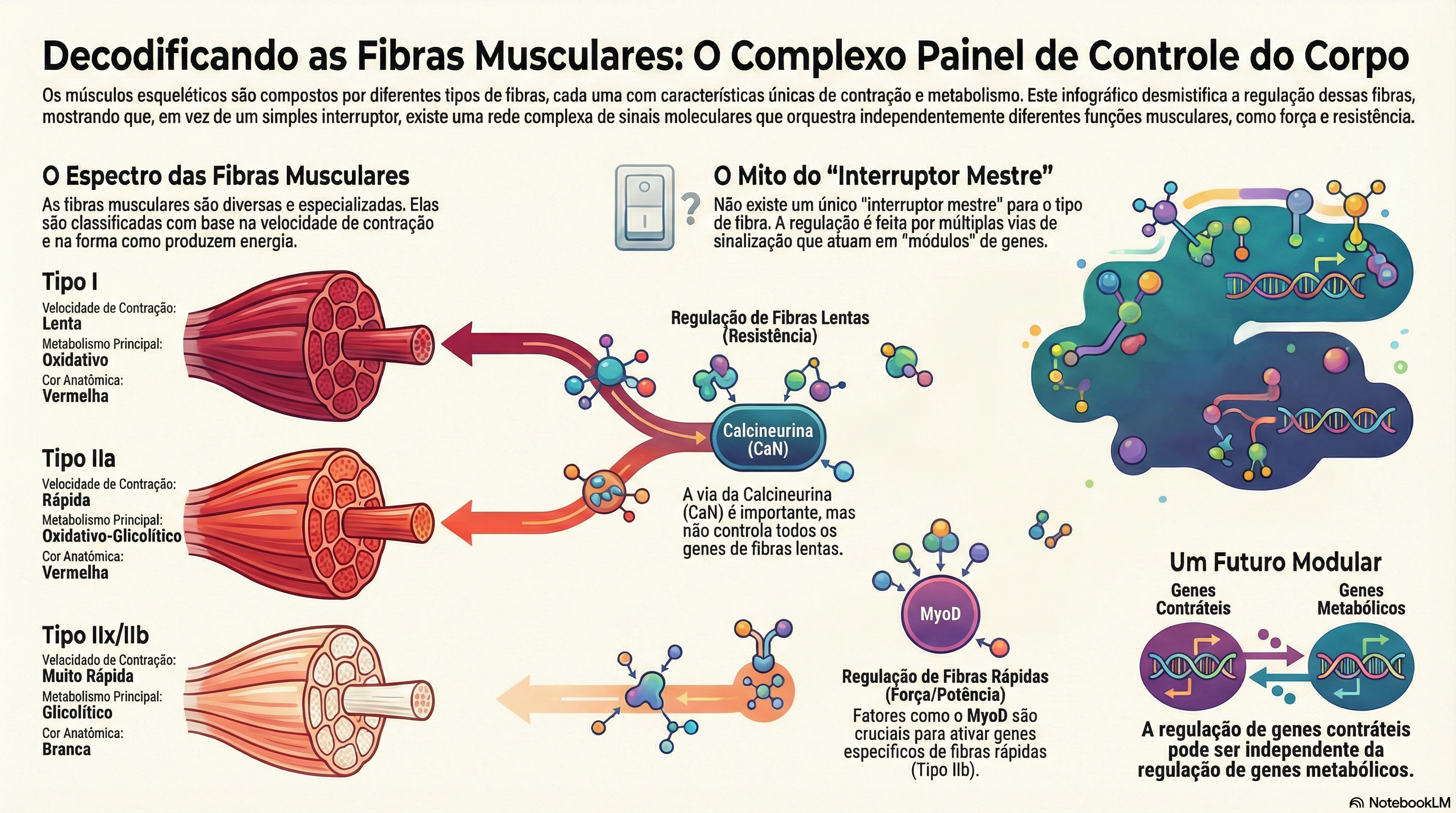
O artigo de apresenta uma revisão crítica sobre como o fenótipo de tipos de fibras musculares esqueléticas é regulado em nível celular e molecular. A tese central é explícita: a expressão fenotípica associada aos “tipos de fibras” não é controlada por um único “interruptor mestre”, mas por múltiplas vias de sinalização e combinações de fatores de transcrição, com impacto parcial e por vezes dissociado sobre diferentes conjuntos funcionais de genes.
Os autores iniciam contextualizando a própria evolução histórica da nomenclatura. A classificação “vermelhas” e “brancas” (século XIX) evoluiu para categorias que combinavam velocidade contrátil e capacidade oxidativa, e, posteriormente, para a classificação baseada em isoformas de miosina de cadeia pesada (MHC): tipo I, IIa, IIx e IIb (com a ressalva de que, em humanos, a expressão de IIb não é típica em músculos locomotores). Ainda assim, o texto enfatiza um ponto metodológico decisivo: a classificação por MHC é, estritamente, uma “classificação por MHC”, e não uma descrição exaustiva do “tipo de fibra” como entidade funcional completa. Isto ocorre porque muitos outros componentes proteicos (enzimas metabólicas, transportadores, proteínas de manejo de cálcio, etc.) podem não variar em paralelo com as isoformas de MHC, e há coexistência de múltiplas isoformas em uma mesma fibra (fibras “híbridas”), o que amplia a heterogeneidade para além de quatro categorias discretas. O artigo sugere, portanto, que uma interpretação mais consistente é modular: “tipos de fibras” seriam combinações de “domínios funcionais” (por exemplo, maquinário contrátil; pool mitocondrial; proteínas de transporte iônico; vias glicolíticas; oxidação de ácidos graxos), que podem ser regulados de forma parcialmente independente.
Na seção sobre sinalização, o artigo usa o debate em torno da calcineurina (CaN) como exemplo didático do problema de se inferir regulação global a partir de um subconjunto de marcadores. A CaN é uma fosfatase sensível a cálcio (Ca²⁺) que, via desfosforilação, promove a translocação nuclear de fatores NFAT (nuclear factor of activated T cells). Uma motivação clássica para envolvê-la na especificação lenta/oxidativa é a capacidade de discriminar padrões de Ca²⁺: sinais sustentados e de baixa amplitude (mais compatíveis com padrões de ativação de fibras lentas) tenderiam a favorecer sua ativação, enquanto picos transitórios de alta amplitude (compatíveis com fibras rápidas) não necessariamente. Estudos citados na revisão mostram translocação nuclear de NFAT sob padrões de estimulação que mimetizam atividade “lenta”, e ausência desse efeito em padrões “rápidos”. Entretanto, o artigo reúne um conjunto de resultados que limita qualquer leitura “monocausal”: (i) a ativação constitutiva de CaN pode aumentar a atividade de promotores associados a genes “lentos” em alguns modelos, mas não ativa todos os genes típicos de fibras lentas; (ii) há evidências de ativação também de promotores “rápidos” por CaN em determinadas condições; (iii) em injeções in vivo, um construto constitutivamente ativo de CaN não necessariamente aumenta a atividade de determinados promotores “lentos”; (iv) intervenções farmacológicas com ciclosporina A (CsA), frequentemente usadas para inferir papel de CaN, podem gerar interpretações ambíguas por efeitos sistêmicos e mitocondriais não específicos, além de variabilidade na inibição efetiva da atividade de CaN entre músculos com perfis distintos de subunidades. Um dado quantitativo destacado é que, em um estudo, CsA elevou a proporção de fibras expressando MHC “rápida” no sóleo de rato de 14% para 31%, mas a revisão ressalta que achados semelhantes não são universalmente reproduzidos, e que a própria eficácia de CsA pode variar entre músculos (dependendo da composição de subunidades e sensibilidade do complexo).
O texto também organiza a CaN como parte de uma lógica combinatória: NFAT frequentemente atua em conjunto com MEF2 (myocyte enhancer factor 2) para transduzir sinais dependentes de Ca²⁺ em mudanças transcricionais. Elementos regulatórios em promotores “lentos”, como o SURE (slow upstream regulatory element) do gene troponina I lenta (TnIs), contêm motivos de ligação para MEF2 em regiões A/T ricas e também sítios relacionados a NFAT/E-box/CACC. A revisão enfatiza que mutações isoladas de um motivo (por exemplo, NFAT) podem ser insuficientes para abolir especificidade ou ativação, sugerindo redundância e necessidade de múltiplos fatores atuando em conjunto. A proposta conceitual é que vias como CaN–NFAT e CaN–MEF2 são relevantes, mas exercem controle sobre subconjuntos de genes e não sobre toda a “identidade” fenotípica de um tipo de fibra.
Outra via examinada envolve CaMK (calcium/calmodulin-dependent protein kinase) e seu potencial papel na regulação de fibras tipo I e no acoplamento com biogênese mitocondrial. Em modelos transgênicos com CaMKIV constitutivamente ativa, há relato de aumento de fibras tipo I no plantaris de 2% para 10%, novamente interpretado pelos autores como um efeito real, porém parcial, insuficiente para sustentar a ideia de “conversão completa” por um único sinal. Ainda nesse eixo aparece PGC-1α (peroxisome proliferator-activated receptor gamma coactivator 1-alpha), coativador transcricional que interage com fatores ligados ao DNA (como NRF-1) e é reiteradamente associado à biogênese mitocondrial. O artigo descreve que PGC-1α pode aumentar marcadores mitocondriais e alguns genes associados a fenótipo mais oxidativo; contudo, mesmo em transgênicos, o aumento de fibras tipo I pode permanecer em torno de ~10% em certos músculos, e a coativação com outras vias (por exemplo, CaN) pode amplificar respostas em promotores específicos como o da mioglobina, dependente de sítio de MEF2. Ainda assim, o texto chama atenção para a extrapolação para humanos: a revisão aponta que, em humanos, treinamento de endurance eleva marcadores de densidade mitocondrial, mas tipicamente não altera a porcentagem de fibras tipo I, o que enfraquece a interpretação de PGC-1α como “determinante principal” de “especificação” de fibra tipo I em sentido amplo; sustenta, ao contrário, a ideia de que módulos (mitocondrial/oxidativo) podem ser regulados sem obrigatoriamente redefinir o módulo contrátil (MHC).
No domínio do metabolismo energético e biogênese mitocondrial, a revisão incorpora também AMPK (AMP-activated protein kinase) como sensor de estresse energético (alteração da razão ATP/AMP) com ligação a NRF-1 e aumento de componentes respiratórios e densidade mitocondrial em condições de ativação crônica. A discussão mantém o mesmo fio: essas vias são plausíveis e suportadas por dados para “pools” mitocondriais, mas não implicam, por si, reprogramação total de todos os atributos de um tipo de fibra.
Para fibras rápidas glicolíticas (IIx/IIb, no contexto de roedores), o artigo detalha elementos regulatórios como FIRE (fast intronic regulatory element) e a relevância de E-box e fatores miogênicos (MyoD, myogenin, Myf-5) no controle de promotores como o de MHC IIb. Um ponto enfatizado é a dependência do contexto: mutações do mesmo tipo de motivo (por exemplo, CArG) podem produzir efeitos opostos em promotores distintos (IIa vs IIx vs IIb), sugerindo que a “mesma sequência” pode ser interpretada de maneira diferente dependendo do conjunto de fatores presentes, interações entre sítios e arquitetura do promotor. A revisão também traz dados em que a ausência de MyoD reduz a indução de MHC IIb em condições de desuso (hindlimb unweighting), mas novamente sem concluir exclusividade, pois ainda há expressão residual em músculos nulos para MyoD. O argumento retorna ao núcleo: a fibra preserva plasticidade justamente porque a regulação é distribuída, redundante e combinatória; um único fator dificilmente seria capaz de governar, com fidelidade e flexibilidade, a expressão coordenada de centenas ou milhares de produtos gênicos.
O artigo inclui ainda uma síntese direcional das adaptações de tipos de fibras ao treinamento e ao desuso. Em roedores, endurance é descrito como aumentando fibras tipo I e IIa e reduzindo IIx/IIb; em humanos, endurance é descrito como não alterando a proporção de tipo I, aumentando IIa e reduzindo IIx (sem IIb). Para treinamento de força, a revisão também indica em humanos um padrão típico de aumento de IIa e redução de IIx, sem mudança relevante em tipo I. A interpretação coerente com toda a revisão é que “mudanças de perfil” frequentemente refletem redistribuições dentro do espectro rápido (especialmente IIx→IIa) e mudanças metabólicas importantes em todos os tipos, sem que isso exija “conversões completas” entre extremos (rápida↔lenta) em humanos.
A conclusão é uma crítica metodológica e conceitual: (1) deve-se evitar inferir “regulação do tipo de fibra” a partir de regulação de um único promotor ou de poucos marcadores; (2) investigações deveriam avaliar se uma via candidata afeta múltiplos módulos (contrátil e metabólico, por exemplo) e não apenas um domínio; (3) a classificação atual baseada em MHC é útil, mas provavelmente insuficiente frente a abordagens “pós-genômicas”. A previsão proposta é que classificações futuras possam se apoiar em assinaturas regulatórias (famílias de genes com regiões regulatórias comuns e responsivas à atividade contrátil) em vez de categorias discretas baseadas apenas em uma proteína estrutural.
Referência: Spangenburg EE, Booth FW. Molecular regulation of individual skeletal muscle fibre types. Acta Physiol Scand. 2003;178:413–424.

O professor que “não está na prática”
23-06-2026


