Musculação na Terceira Idade: Força, Função e Qualidade de Vida
O “treinamento híbrido” e o desserviço pedagógico da reinvenção de conceitos já consolidados
Cronograma dos Cursos 2025
A Faixa Preta e o Doutorado
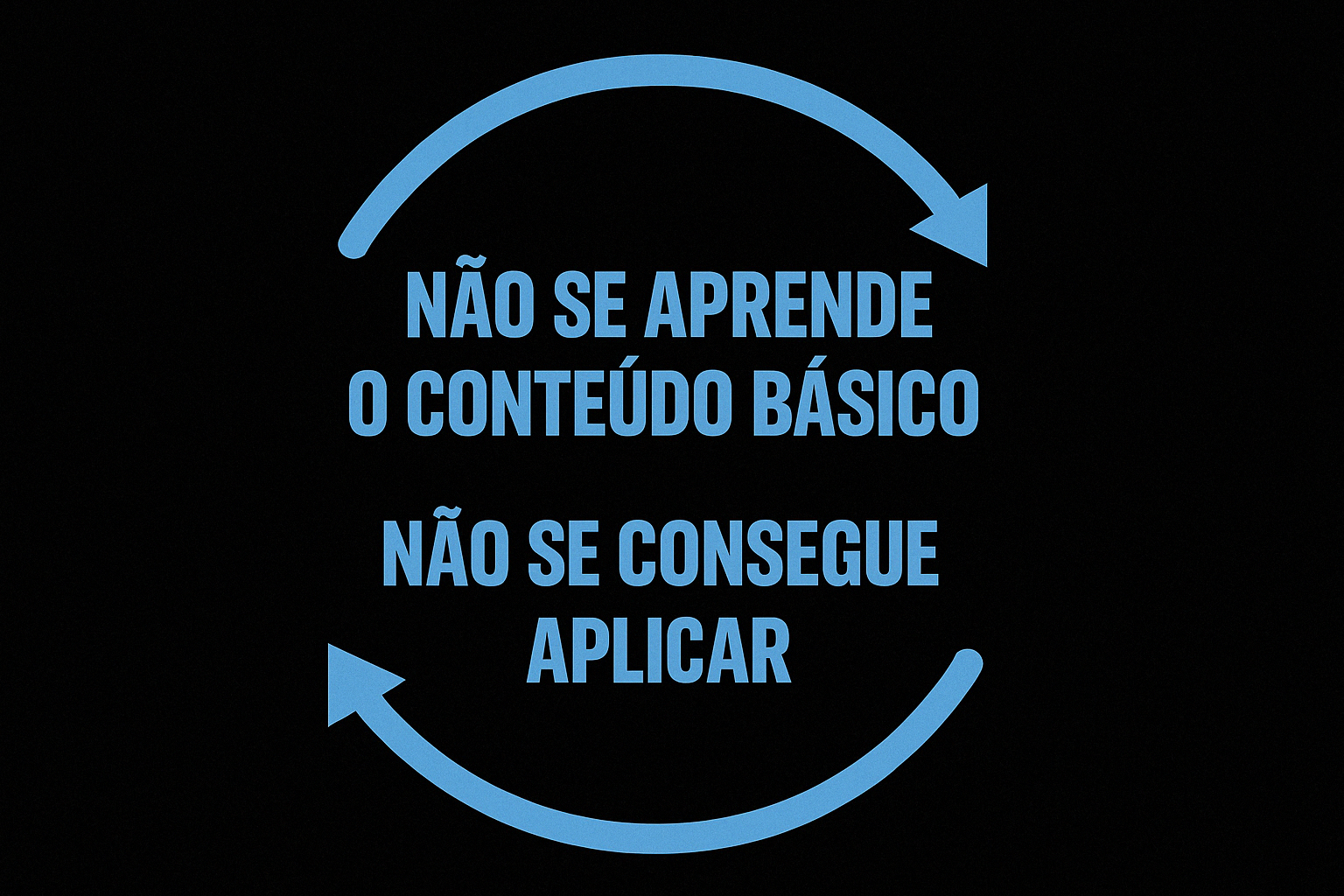
O ciclo da superficialidade no ensino das ciências do esporte: quando a busca por aplicação imediata compromete a formação científica.
Copyright © 2021